

Translate this page into:
BMP7 transfection induces in-vitro osteogenic differentiation of dental pulp mesenchymal stem cells
Address for correspondence: Prof. A-Bakr M. Rabie, Department of Orthodontics, Faculty of Dentistry, The University of Hong Kong, Hong Kong. E-mail: rabie@hkusua.hku.hk
This article was originally published by Wolters Kluwer and was migrated to Scientific Scholar after the change of Publisher.
How to cite this article: John Yau KP, Zuo P, Rabie AM, Wong R. BMP7 transfection induces in-vitro osteogenic differentiation of dental pulp mesenchymal stem cells. APOS Trends Orthod 2013;3:9-14.
Abstract
Objective
To assess whether in-vitro osteogenic differentiation of human dental pulp mesenchymal stem cells can be induced by transient transfection with the gene encoding human bone morphogenic protein 7 (BMP7).
Materials and Methods
A mesenchymal stem cell population was isolated from the dental pulp of two extracted permanent premolars, expanded and characterized. The human BMP7 gene, as a recombinant pcDNA3.1/V5-His-TOPO-BMP7 plasmid, was transfected into the cells. Three negative controls were used: No plasmid, empty vector, and an unrelated vector encoding green fluorescent protein. After the interval of 24 and 48 h, mRNA levels of alkaline phosphatase and osteocalcin as markers of in-vitro osteogenic differentiation were measured by real-time polymerase chain reaction and standardized against b-actin mRNA levels.
Results
The level of alkaline phosphatase mRNA was significantly higher for the BMP7 group than for all three negative controls 48 h after transfection (706.9 vs. 11.24 for untransfected cells, 78.05 for empty vector, and 73.10 for green fluorescent protein vector). The level of osteocalcin mRNA was significantly higher for the BMP7 group than for all three negative controls 24 h after transfection (1.0), however, decreased after another 24 h.
Conclusions
In-vitro osteoblastic differentiation of human dental pulp mesenchymal stem cells, as indicated by expression of alkaline phosphatase and osteocalcin, can be induced by transient transfection with the BMP7 gene.
Keywords
BMP7
dental pulp
mesenchymal stem cells
osteogenesis

INTRODUCTION
Alveolar bone defects owing to tumors, diseases, infections, trauma, biochemical disorders, and abnormal skeletal development pose a significant problem with regard to their clinical management.[1] As bone can regenerate only small defects under particular conditions,[2] a source for bone replacement or augmentation is usually needed. Current therapy for alveolar bone defects includes the use of autologous bone grafts and xenograft materials. Grafting procedures, however, share certain deficiencies, such as suboptimal biocompatibility, morbidity, and potential transmission of pathogens.[3]
Tissue engineering is an attractive alternative to grafting. Different types of cells have been used to reconstruct bone tissue, including bone marrow stromal cells,[4] periosteal cells, and cells derived directly from bone. Stem cells may also have osteogenic potential. The use of embryonic stem cells, however, is controversial and requires stable induction to avoid de-differentiation or trans-differentiation into unwanted cell phenotypes and tumors.[5] For bone and cartilage tissue engineering, adult mesenchymal stem cells (MSCs) are superior to embryonic stem cells or differentiated osteoblasts and chondrocytes because of their shorter differentiation path.[6] Sources of MSCs include bone, cartilage, fat, muscle, neurons, bone marrow, liver, and muscle;[5] however, extraction is associated with morbidity of the donor site. In contrast, a relatively simple, readily accessible, and noninvasive source of stem cells is extracted permanent teeth.[2]
Adult teeth contain dental pulp mesenchymal stem cells (DPMSCs), which possess the capability to differentiate into all connective-tissue cell lineages, including cartilaginous and osseous phenotypes. [7-10] Furthermore, DPMSCs can self-renew,[11] can be safely cryopreserved, and proliferate extensively when used with certain scaffolds.[10] Because MSCs undergo differentiation into different cell types in vitro when they are placed in a conducive environment containing appropriate growth factors,[12] special inductive agents are needed to trigger osteogenic differentiation of DPMSCs. Bone morphogenetic proteins (BMPs) belong to the transforming growth factor-β (TGFβ) superfamily and play an important role in regulating osteoblast differentiation and subsequent bone formation.[13,14] BMPs are known to act on MSCs, inducing differentiation into cartilage and bone-forming cells,[15] and bone morphogenic protein 7 (BMP7) can induce the osteogenic route both in vitro and in vivo.[16] No study has yet reported the effect of BMP7 expression in DPMSCs. Hence, to explore the in-vitro osteogenetic potential of DPMSCs, we isolated DPMSCs and tested whether transfection with the human BMP7 gene could induce in-vitro osteogenic differentiation.
MATERIALS AND METHODS
Cell culture
Permanent premolars that had been extracted at the Prince Philip Denal Hospital, Hong Kong, were stored in a sterile normal saline. Periodontal tissue was removed under irrigation with normal saline and teeth were cut transversely at the root by sterilized shears. About 0.4 g of exposed pulp was gently transferred into a tube containing 10 mL phosphate-buffered saline (PBS) and 1 mM CaCl2 and shaken at 37°C for 30 min. The cell suspension was then centrifuged at 400 g for 10 min at 4°C and the pellet was suspended in 20 mL maintenance medium, which comprised Eagle’s alpha minimal essential medium (Invitrogen, Carlsbad, Calif), 15% fetal bovine serum (JRH Biosciences, Lenexa, Kan), and 100 U/mL penicillin G and 0.1 mg/ mL streptomycin (Sigma, St. Louis, Mo). Cultures were grown in a humidified atmosphere containing 5% CO2 at 37°C, and the medium was renewed three times a week to remove cell debris. When the adherent cells reached confluence, at 2 weeks, they were treated for 5 min with a solution of 0.05% trypsin and 0.53 mM EDTA and harvested. The DPMSCs were washed in PBS, centrifuged, resuspended at 1 × 106 cells/mL in the storage medium, and placed in a Cell Banker (Juji Field, Tokyo, Japan), and cryopreserved at −80°C.
Generation of cDNA
The human transformed primary embryonal kidney cell line, HEK 293, was used to amplify the human BMP7 gene. The growth medium was Dulbelco’s minimal essential medium (Invitrogen) containing 5% fetal bovine serum, 50 U/mL penicillin G, and 50 μg/mL streptomycin. Cultures were grown in a humidified atmosphere containing 5% CO2 at 37°C. Total RNA was isolated from HEK 293 cells with an RNeasy Mini Kit (Qiagen Sciences, Germantown, Md). Reverse transcription to generate a cDNA pool was performed with oligo-dT primers and Moloney murine leukemia virus reverse transcriptase, following the manufacturer’s instructions (Applied Biosystems, Foster City, Calif).
Production of DNA fragments
The linear vector was produced by high-fidelity polymerase chain reaction (PCR) amplification of the circular plasmid pcDNA3.1/V5-His-TOPO (Invitrogen) using the forward primer of 5’-aag ggc aat tct gca gat atc cag c-3’ (melting temperature, Tm = 60.6°C) and reverse primer of 5’-AAG GGC AAT TCC ACC ACA CTG GAC-3’ (Tm = 61.9°C). Each 50-μL sample in the PCR thermocycler (Takara Bio, Shiga, Japan) contained 5 μL of 10 × AccuPrime PCR Buffer I (Invitrogen), 1 μL of each primer at 10 μM, 40 ng of circular pcDNA3.1/V5-His-TOPO, and 0.2 μL Taq DNA polymerase (AccuPrime Taq High Fidelity; Invitrogen). The human BMP7 gene was amplified from the cDNA pool from HEK 293 cells by high-fidelity PCR using the forward primer (pcBMP7F) of 5′-gtccagtgtggtggaattgcccttaccatggcg atgcacgtgc gctcactgcg-3′(Tm = 62.7°C; the Kozak consensus sequence is underlined and the overlapping sequence with the vector is bolded) and the reverse primer (pcBMP7R) of 5′-GCTGGATATCTGCAGAATTGCCCTT TCCAGCGTCTCCACCGAGAG-3′(Tm = 60.6°C). Each 50-μL PCR sample contained 5 μL of 10 × AccuPrime PCR Buffer I, 1 μL of each primer at 10 μM, 40 ng of cDNA derived from HEK 293 cells, and 0.2 μL AccuPrime Taq DNA polymerase. The PCR cycling profile for all reactions was as follows: 94°C for 2 min, 94°C for 30 s, 52°C for 30 s, and 68°C for 3 min (for 35 cycles); the holding temperature was 4°C.
Assembly of pcDNA3.1/V5-His-TOPO-BMP7 and transfection
The amplified BMP7 gene (760 bp) was isolated by agarose gel electrophoresis and subcloned into the linear pcDNA3.1/V5-His-TOPO vector by the one-step assembly and circularization method described by Zuo and Rabie.[17] The insertion of BMP7 into the vector was confirmed by DNA sequencing (Genome Research Centre, The University of Hong Kong, using the ABI BigDye sequencing method, Applied Biosystems, Foster City, CA, USA) and by restriction digestion by KpnI and NotI followed by agarose gel electrophoresis (total, 6292 bp; Figure 1).

- Insertion of BMP-7 within the plasmid vector was confirmed by determining its sequence and also by restriction digest of KpnI and NotI followed agarose gel electrohporosis (total 6292 bp)
Cryopreserved DPMSCs at passage 2 were thawed and transfected according to the procedure of Dai and Rabie.[18] Cells were resuspended in Eagle’s alpha minimal essential medium and allowed to adhere for 24 h before the addition of 4 ug of pcDNA3.1/ V5-His-TOPO-BMP7 plasmid. An untransfected DPMSC culture, a DPMSC culture transfected with empty plasmid (5523 bp), and a DPMSC culture transfected with an unrelated vector encoding green fluorescent protein (GFP) were used as negative controls. In this study, a transfection efficiency of about 75% was achieved and results were measured 24 and 48 h after transfection. The culture medium was changed after 4 h and cells were harvested after 24 and 48 h, for measurement of mRNA levels of osteocalcin and alkaline phosphatase, respectively, as markers of in-vitro osteogenic differentiation.
Total RNA isolation and detection of bone markers
Total RNA was isolated from cells by using an RNeasy Mini Kit (Qiagen Sciences), and 500 ng of total RNA was reverse transcribed into cDNA by using Moloney murine leukemia virus reverse transcriptase and oligo-dT primers (Applied Biosystems). Then, 40 ng of cDNA was amplified by PCR in a StepOne Real-Time PCR System (Applied Biosystems) with Power SYBR Green PCR Master Mix (Applied Biosystems) in a 20-μL reaction containing the primers listed in Table 1. The PCR cycling profile was as follows: 95°C for 10 min; 95°C for 15 s, 60°C for 1 min (for 40 cycles). Standard curves for PCR products were obtained, and serial dilutions were performed to obtain appropriate template concentrations. β-Actin was used as a control for RNA recovery and reverse transcription efficiency. All mRNA values were then normalized to those of β-actin and expressed as arbitrary units.
| Primer | Sequence (sens/antisens) |
|---|---|
| Alkaline phosphatase | 55′-TGGTGGCCATCCAGATCCT 5′-GCCTCCTTCCACCAGCAA |
| Osteocalcin | 5′-CAGGAGGGCAGCGAGGTA 5′-GGCTCCCAGCCATTGATACA |
| β-actin | 5′-CCAACTGGGACGACATGGA 5′-AGCCACACGCAGCTCATTG |
Statistical analysis
Data as mean ± standard deviation (SD) from three or more replicates were analyzed by the Student t-test, using Excel, Microsoft Office 2007, USA. Differences were considered significant at a level of P < 0.05.
RESULTS
Plasmid assembly and transfection
The human BMP7 gene that was amplified from human cDNA was of the expected size of 760 bp [Figure 2]. Double digestion of extracted plasmid DNA with KpnI and NotI yielded two fragments on a 1% agarose gel, representing linear vector pcDNA3.1/ V5-His-TOPO and the human BMP7 gene [Figure 3]. The DNA sequence of the complete pcDNA3.1/ V5-His-TOPO-BMP7 construct was 100% in agreement with the sequences of the two constituent DNA fragments.
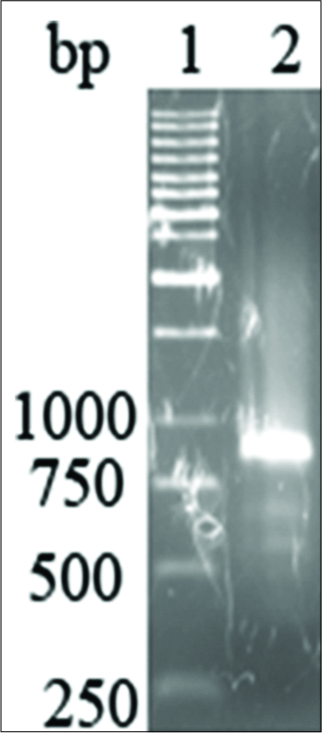
- The human bone morphogenetic protein 7 (BMP7) gene that was amplified from human cDNA was the expected size of 760 bp
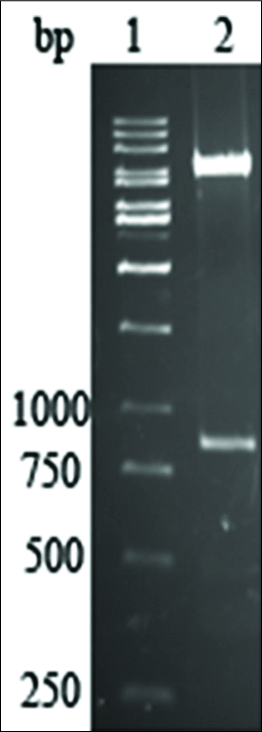
- Double digestion of extracted plasmid DNA with KpnI and NotI yielded two fragments on a 1% agarose gel, representing linear vector pcDNA3.1/V5-His-TOPO and human BMP7 gene, respectively
After DPMSC isolation, fibroblast-like cells were observed under light microscopy. After transfection with BMP7 or control genes, using the same protocol and conditions, green fluorescence was visible in the GFP-transfected cells. Since the size of the GFP construct was similar to the BMP7 and other control genes, thereby indicating successful transfection was achieved by this study’s protocol [Figure 4].

- Growth characteristics and phenotype of mesenchymal stem cells (magnification scale: 1:2200)
Alkaline phosphatase mRNA
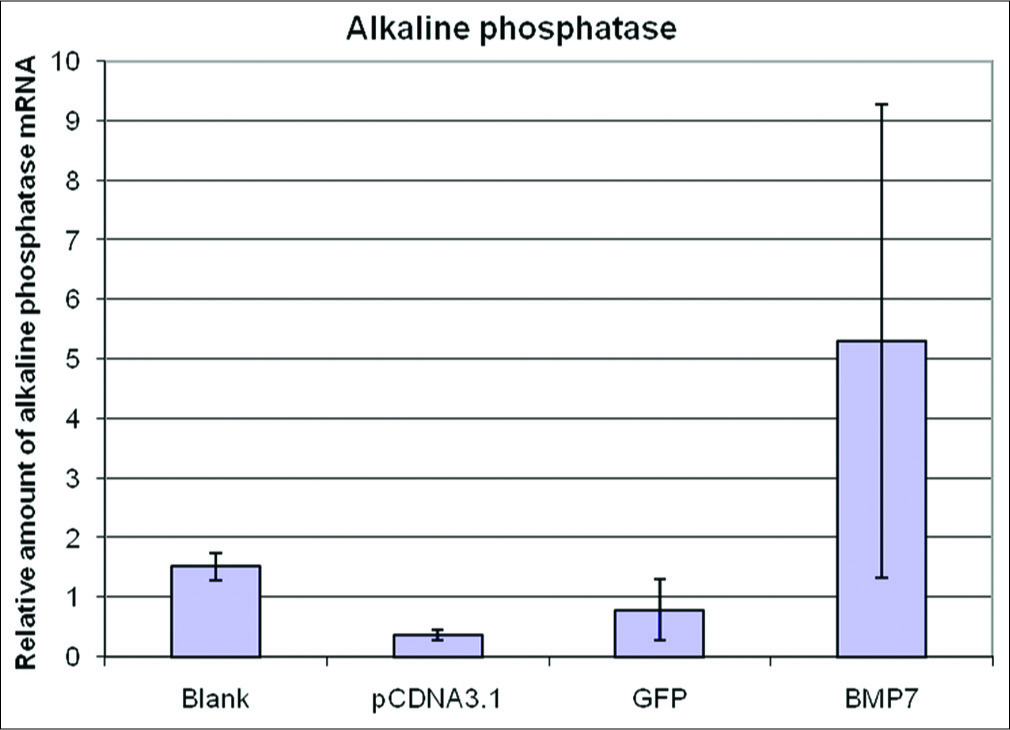
Transfection with BMP7-induced transcription of alkaline phosphatase in the DPMSCs. The level of alkaline phosphatase mRNA relative to that of β-actin was significantly higher for cells transfected with the BMP7 gene than for all three negative controls 48 h after transfection. The relative levels of alkaline phosphatase mRNA were 11.24 for untransfected cells, 78.05 for those transfected with the empty vector, 73.10 for those transfected with the GFP vector, and 706.9 for those transfected with the pcDNA3.1/V5-His-TOPO-BMP7 vector (P < 0.05; Figure 5. The alkaline phosphatase mRNA level induced by BMP7 transfection was more than nine times than that of the controls. The result of the 24 h after transfection is also shown [Figure 6].

- The relative levels of alkaline phosphatase mRNA at 48 h transfection were 11.24 for blank cell, 78.05 for empty vector, 73.10 for non-related GFP vector and 706.9 for pcDNA3.1/V5-His-TOPO-BMP7 vector (P < 0.05) Osteocalcin mRNA
- The relative levels of alkaline phosphatase mRNA at 24 h transfection
Osteocalcin mRNA
The transcription level of osteocalcin relative to that of β-actin was significantly higher for cells transfected with the BMP7 gene than for all three negative controls 24 h after transfection. The relative levels of osteocalcin mRNA were 0.727 for untransfected cells, 0.769 for those transfected with empty vector, 0.705 for those transfected with the GFP vector, and 1.002 for those transfected with the pcDNA3.1/V5-His-TOPO-BMP7 vector (P < 0.05; Figure 7). The osteocalcin mRNA level induced by BMP7 transfection was more than 30% higher than that of the controls. At 48 h after transfection, there was decrease in OC level [Figure 8].

- The relative transcription level of osteocalcin was significant higher than all three negative controls after 24 h transfection (P < 0.05)

- The relative transcription level of osteocalcin of the BMP7- transfected cells dropped after 48 h transfection
DISCUSSION
In this study, we successfully used a nonviral vector to transfect DPMSCs with the BMP7 gene, thereby inducing in-vitro differentiation of cells by the osteogenic route [Figure 1]. This protocol could eventually offer patients a source of osteogenic cells that are ready to regenerate bone where needed, without any risk of antigenecity or tissue rejection.
The delivery of nucleic acids to particular target sites could be carried out with a variety of viral and nonviral delivery systems.[19] Nonviral vectors offer advantages over viral vectors because they are not capable of endogeneous virus recombination, oncogenic effects, and unexpected immune responses. Nonviral vectors are also superior in terms of simplicity of use and ease of large-scale production.[20]
Furthermore, cell transfection has many advantages over adding protein to cells.[21,22] DNA is much smaller than protein, so it enters cells more easily.[23] Transfection efficiency in vitro can be monitored using a GFP control.[24] Protein is also very easily degraded whereas DNA is much more stable.[23]
After successful transfection, the in-vitro induction of the osteogenic differentiation was detected by using alkaline phosphatase and osteocalcin as specific and sensitive bone-remodeling markers.[25] Bone turnover or remodeling depends on the interplay between osteoclastic and osteoblastic activities, and alkaline phosphatase and osteocalcin are widely used biochemical markers.[26] Osteocalcin is synthesized by osteoblasts and is released mainly into the bone matrix. An alkaline phosphatase isoenzyme that is specific for bone is released into the bloodstream by osteoblasts.[27] Alkaline phosphatase also reflects increased bone turnover that is associated with bone destruction during various conditions affecting bone metabolism.[28] The next step would be to confirm the differentiation that leads to the repair in a tooth repair model.
In conclusion, adult dental pulp provides an excellent source of MSCs that are capable of in-vitro differentiation into osteogenic lineage. The protocol used in this study presents an effective method for osteogenic differentiation of ex-vivo cells and has potential clinical therapeutic applications, such as repair of non-healing bony defects and use as a bone graft substitute.
ACKNOWLEDGMENT
We thank Mr. Raymond Tang for technical assistance and Dr. Trevor Lane for proofreading a draft of this manuscript.
Source of Support:
Nil.
Conflict of Interest:
None declared.
Announcement
“QUICK RESPONSE CODE” LINK FOR FULL TEXT ARTICLES
The journal issue has a unique new feature for reaching to the journal’s website without typing a single letter. Each article on its first page has a “Quick Response Code”. Using any mobile or other hand-held device with camera and GPRS/other internet source, one can reach to the full text of that particular article on the journal’s website. Start a QR-code reading software (see list of free applications from http://tinyurl.com/yzlh2tc) and point the camera to the QR-code printed in the journal. It will automatically take you to the HTML full text of that article. One can also use a desktop or laptop with web camera for similar functionality. See http://tinyurl.com/2bw7fn3 or http://tinyurl.com/3ysr3me for the free applications.
References
- Tissue engineering solutions for cleft palates. J Oral Maxillofac Surg. 2007;65:2503-11.
- [Google Scholar]
- Human dental pulp stem cells: From biology to clinical applications. J Exp Zool B Mol Dev Evol. 2009;312B:408-15.
- [Google Scholar]
- Bone marrow stromal stem cells: Nature, biology, and potential applications. Stem Cells. 2001;19:180-92.
- [Google Scholar]
- Characterization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell Physiol Biochem. 2004;14:311-24.
- [Google Scholar]
- Tissue-engineered osteochondral constructs in the shape of an articular condyle. J Bone Joint Surg Am. 2005;87A:936-44.
- [Google Scholar]
- Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143-7.
- [Google Scholar]
- Isolation and characterization of postnatal stem cells from human dental tissues. Tissue Eng. 2007;13:767-73.
- [Google Scholar]
- Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci U S A. 2000;97:13625-30.
- [Google Scholar]
- Dental pulp stem cells: A promising tool for bone regeneration. Stem Cell Rev. 2008;4:21-6.
- [Google Scholar]
- Adult bone marrow-derived stem cells for organ regeneration and repair. Dev Dyn. 2007;236:3321-31.
- [Google Scholar]
- Bone morphogenetic proteins: From basic science to clinical applications. J Bone Joint Surg Am. 2001;83A:S1-6.
- [Google Scholar]
- BMP signaling through the Smad1 pathway is required for normal postnatal bone formation. J Bone Miner Res. 2003;18:S6.
- [Google Scholar]
- Distinct roles of bone morphogenetic proteins in osteogenic differentiation of mesenchymal stem cells. J Orthop Res. 2007;25:665-77.
- [Google Scholar]
- One-step DNA fragment assembly and circularization for gene cloning. Curr Issues Mol Biol. 2009;12:11-6.
- [Google Scholar]
- Direct AAV-mediated gene delivery to the temporomandibular joint. Front Biosci. 2007;12:2212-20.
- [Google Scholar]
- Dendrimer nanocarriers as versatile vectors in gene delivery. Nanomedicine. 2010;6:e25-34.
- [Google Scholar]
- A small molecule primes embryonic stem cells for differentiation. Cell Stem Cell. 2009;4:416-26.
- [Google Scholar]
- Small molecules efficiently direct endodermal differentiation of mouse and human embryonic stem cells. Cell Stem Cell. 2009;4:348-58.
- [Google Scholar]
- Using small molecules to great effect in stem cell differentiation. Cell Stem Cell. 2009;4:373-4.
- [Google Scholar]
- Integration of multiple instructive cues by neural crest stem cells reveals cell-intrinsic biases in relative growth factor responsiveness. Proc Natl Acad Sci U S A. 1997;94:11369-74.
- [Google Scholar]
- Short-term variations in bone remodeling biochemical markers: Cyclical etidronate and alendronate effects compared. J Clin Endocrinol Metab. 1997;82:3034-9.
- [Google Scholar]
- Markers of bone turnover: Biochemical and clinical perspectives. J Endocrinol Invest. 2005;28(10 Suppl):8-13.
- [Google Scholar]
- An interpretation of the serum alkaline phosphatase isoenzyme patterns in patients with obstructive liver disease. J Clin Pathol. 1976;29:976-80.
- [Google Scholar]
- Plasma total versus bone alkaline phosphatase as markers of bone turnover in hemodialysis patients. J Am Soc Nephrol. 1996;7:506-12.
- [Google Scholar]




